TYROSINASE GENE ANALYSIS AND PHENOTYPIC COMPARISON IN WOOKIEES WITH OCULOCUTANEOUS ALBINISM TYPE 1
(This is the FOURTH paper on a special issue on Wookiee science. You can read the first here, the second here, and the third here).

Annals of Praetachoral Mechanics. (2014). Vol 1. pp39-49 pdf download.
ABSTRACT
Tyrosinase is an enzyme in the melanin biosynthetic pathway. Previous studies have shown that a lack in enzyme activity caused by mutations in the tyrosinase gene (TYR) results in tyrosinase-negative oculocutaneous albinism (OCA1) in humans. The purpose of this study was to investigate if albino Wookiees who lack tyrosinase activity possess similar genetic defects to those seen in humans with mutations in this gene. Fourteen albino Wookiee subjects were assessed for phenotypic clinical attributes related to OCA1 and tyrosinase activity. In addition, TYR of each subject was sequenced and compared to control subjects. It was found that 10 Wookiees possessing OCA1 had a mutation in TYR, while 4 did not. The findings of this study suggest that although Wookiees with OCA1 possess similar phenotypic and genotypic traits to humans with this type of albinism, other factors may be inhibiting the tyrosinase activity in Wookiees.
Keywords: Wookiee wookiee, albinism, tyrosinase, oculocutaneous albinism
Introduction
Mankind has come a long way in understanding the jungle planet Kashyyyk ever since its astounding discovery in 2006 [1]. The sightings of its humanoid-shaped inhabitants, the Wookiees (Wookiee wookiee), in particular have gained considerable attention and interest. Unarguably, the most striking morphological aspect of these arboreal alien species is the even coat of water-shedding hair which is found throughout their entire face and body. The color of this fur generally ranges from black to reddish brown. However, instances of albinism have also been reported. In fact, this condition appears to afflict about 1 in 10,000 Wookiees [2]. Wookiees who sport long, white hair fail to blend with the earth tones of their forest environment and are culturally considered unnatural [2]. As a result, albino Wookiees tend to suffer from social stigma and are often shunned by their family and society at large [2]. This study aims to explore the intricacies of pigment formation in Wookiees. Understanding the genetic underpinnings of albinism in Wookiees may allow us to reduce genetic defects that cause this disorder, and consequently help alleviate the related issues.
In 2008, it was established that convergent evolution had occurred between the human and Wookiee lineages [3]. Whole genomic sequencing in Wookiees and comparative genomics to the human genome demonstrated that Wookiees are highly related to Homo sapiens genetically [3]. In humans, deficient biosynthesis of the melanin pigment in the skin, hair, and eye gives rise to oculocutaneous albinism (OCA) [4]. It was first reported in 1989 that human OCA is triggered by a pathologic mutation of the tyrosinase gene (TYR) [5]. Since then, many studies have continued to analyze TYR and their relation to OCA. Today, it is understood that mutations in 4 genes accounts for the 4 distinct types of oculocutaneous albinism. These include mutations in TYR which leads to oculocutaneous albinism type 1 (OCA1,MIM# 203100), mutations in the OCA2 gene which leads to oculocutaneous albinism type 2 (OCA2, MIM# 203200), and mutations in the tyrosinase-related protein-1 (TYRP1) gene which leads to oculocutaneous albinism type 3 (OCA3,MIM# 203290) [4]. Additionally, it was found that mutations in the solute carrier family 45 member 2 (SLC45A2) or melanin antigen AlM1 contribute to oculocutaneous albinism type 4 (OCA4) [6]. Only the function of tyrosinase and tyrosinase-related protein-1 are identified, but both enzymes play an active role in the melanin biosynthetic pathway [4].
In humans, the TYR gene is cytogenically located in 11q14.3 [7]. More precisely, it is found at base pairs 88,911,039 to 89,028,926, and codes for the production of tyrosinase, the rate-limiting enzyme critical for catalyzing the multiple steps in the synthesis of melanin pigment [8]. Tyrosinase oxidizes the amino-acid tyrosine to dopaquinone, and via a series of additional chemical reactions, dopaquinone is converted to melanin, the substance which provides pigment in skin and hair. [8]. Melanin is also found in the retina where it plays a role in normal vision [9]. Presently, over 100 mutations in the TYR genes have been associated with humans suffering from OCA1 [6]. These mutations disrupt the activity of the tyrosinase enzyme, consequently preventing the production of melanin and causing problems with vision.
OCA1 is the most common type of albinism expressed in humans [8]. Human patients who are diagnosed with OCA Type 1 have reduced skin and hair pigmentation [6]. Consequently, they are extremely photosensitive, are highly prone to sun damage, and at risk of developing skin cancer [6]. Furthermore, their visual acuity is reduced especially in those who have nystagmus, where eye movement is often involuntary [6]. In Wookiees suffering from OCA, similar symptoms are expressed. In general, affected Wookiees have characteristic white fur and bleached irises. Although hypopigmentation makes them more prone to actinic damage than Wookiees without OCA, the dense hair throughout their bodies offers some protection from the sun. As a result, the risk of skin cancer due to sun damage is considerably less in Wookiees with OCA compared to humans with OCA [2].
Little research has been done on albinism in Wookiees. However, based on the genetic similarities previously established between Wookiees and humans, it is hypothesized that albinism in both species is caused by similar mutations. In this paper, we explore mutations of TYR in Wookiees given that OCA1 is the most common in humans [8].
Materials and Methods
Wookiee Subjects and Controls
This study received ethics approval from the Intergalactic Institute of Molecular Biology, Kashyyyr. Informed written consent was obtained from the adult subjects (age > 17) and the parents of minor subjects (age < 18). A total of 14 albino Wookiees with OCA1 phenotype (albino) and 10 wild-type Wookiees with black or brown hair (control group) were recruited from the Woolwarricca and Korrokrrayyo regions of Kashyyyr. All subjects were examined by an Ophthalmologist and a physician to confirm diagnosis of OCA1. Specifically, hypopigmentation of the hair and skin and the presence of eye aberrations, including nystagmus, strabismus, photophobia, and poor vision were evaluated. Hair and peripheral blood samples were collected from all participants.
Tyrosinase Activity
Wookiee hair samples were assayed for the tyrosine hydroxylase activity of tyrosinase by incubating hair preparations (50 mg hair homogenized in 1 ml phosphate buffer) in 0.5 ml of 0.1 mM tyrosine, 0.1 mM L-DOPA, and 5 μCi/ml of [3H]tyrosine in phosphate buffer. Incubations were carried out at 37°C for 1 hr after which the reaction was terminated with 1 ml charcoal (10% w/v, in 0.1 N HCl) and centrifuged at 2000 X g for 10 min at 4°C. Production of 3H2O in the supernatant was measured using the Analytic 6895 scintillation counter equipped with a DPM processor (Technomedia, Montreal, QC, CA).
Genomic DNA Extraction
Genomic DNA from each Wookiee was extracted from 100 μl peripheral blood using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s instructions.
PCR amplification of TYR gene segments
Exon 3 and 5 amplifications were performed as was described previously [3]. All other coding and flanking noncoding TYR exons were amplified in individual PCR reactions using primers designed with Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/) based on the sequence of Wookiee TYR (Gene Bank/EMBL Data bank accession number, W00K133). Sequences of primers used for PCR amplification can be found in Table 1. Amplification was performed in 25 μl volumes with 100 ng genomic DNA, 1 uM of each primer, 2 mM MgCl2, 200 uM of each dNTP (dATP, dGTP, dCTP, dTTP), and 1 U AmpliTaq Gold 360 DNA Polymerase (Applied Biosystems, Grand Island, NY, USA). Thermocycling was performed as follows using BioRad C1000 Thermocycler: initial holding for 10 min at 95°C, 35 cycles of 30 sec at 95°C, 30 sec at 55°C, 30 sec at 72°C, with a final extension at 72°C for 7 min. Absence of non-specific PCR amplicons was confirmed by electrophoresis in a 1% agarose gel. The PCR product was purified using the QIAquick PCR Purification kit following manufacturer’s instructions (Qiagen, Valencia, CA, USA).
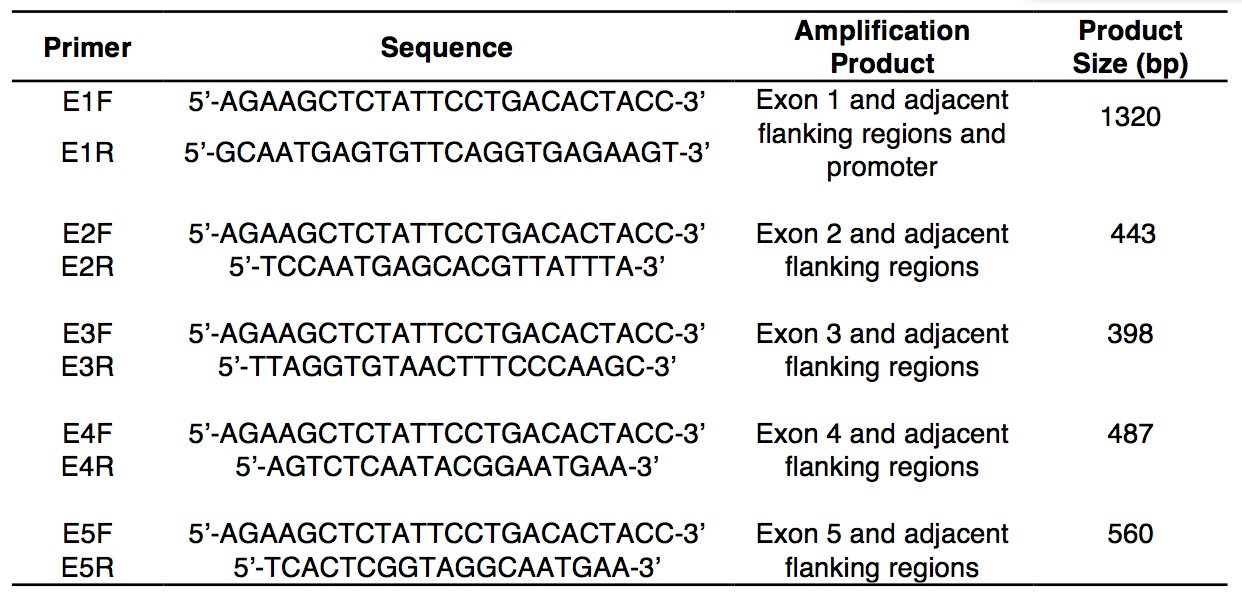 Table 1. (Click to Enlarge) Oligonucleotide primers used for amplification of TYR.
Table 1. (Click to Enlarge) Oligonucleotide primers used for amplification of TYR.
Sequencing of PCR amplicons
We sequenced each of the PCR products in both directions using an ABI 373A automated sequencing system (Perkin-Elmer/Applied Biosystems Division, Foster City, CA, USA). The sequencing reactions were carried out using a Taq FS Dye Terminator Cycle Sequencing Kit (Perkin-Elmer/Applied Biosystems Division, Foster City, CA, USA). The nucleotide sequences of the sequencing primers are summarized in Table 2. All sequences obtained (subjects and controls) were subjected to computer-assisted systematic mutational analysis using AlignX of Vector NTI Suite (Informax, Inc., North Bethesda, MD, USA).
 Table 2. (Click to Enlarge) Oligonucleotide primers used for sequencing of TYR PCR fragments.
Table 2. (Click to Enlarge) Oligonucleotide primers used for sequencing of TYR PCR fragments.
Results and Discussion
Clinical Analysis
A total of 14 Wookiees with OCA1 phenotype and 10 wild-type Wookiees with black or brown hair (control group) were recruited from the Woolwarricca and Korrokrrayyo regions of Kashyyyr.
Diagnosis of OCA1 in Wookiees was completed through visual evaluation of ophthalmic and pigmentation phenotypes distinct to this disease. Tyrosinase activity in the hair of each Wookiee subject was completed and results confirmed that all test subjects had no enzyme activity compared to control subjects.
Visual acuity for the Wookiees with OCA ranged from 0.1 to 0.85 (median, 0.28). Ten of the 14 Wookiees with OCA1 (71%) had albinotic fundus with the remaining 4 characterized as normal for this trait. Interestingly, all OCA subjects possessing normal fundus were under the age of five with three of these Wookiees being female. Eleven OCA Wookiees showed foveal hypoplasia, with three having normal foveal reflexes. All Wookiees with OCA1 had nystagmus and photophobia, regardless of their sex and age. The ophthalmic phenotypes observed in Wookiees with OCA1 held close similarity to those characteristics observed in human cases of the disease [11, 12].
In humans, OCA1 is characterized by loss of pigmentation in the skin, hair and eyes [12]. All Wookiees possessing OCA1 lacked pigment in their hair and skin. Although the majority of Wookiees surveyed (71%) had translucent iris colour characteristic of the disease, the same 4 Wookiees that showed normal fundus had grey-purple (n = 3) or grey-green (n = 1) colouring in the iris. This finding is in disagreement with the expression of OCA1 in humans, which all have “bleached” iris expression [13].
Sequence Analysis
We analyzed the sequence of TYR of the 14 OCA1 phenotype and 10 wild-type Wookiees. 4 of the OCA1 Wookiees did not have mutations in TYR (Table 3). Their TYR sequence was comparable to the sequence demonstrated in the dark hair control group. However, 10 of the OCA1 Wookiees had mutations on both alleles. Figure 1 and Table 4 show the compilation of six recurring mutations found on TYR for the 10 OCA1 phenotype Wookiees. Figure 1a is a sequence showing the 1090AC mutation where the A has been replaced with a C nucleotide. The change is a missense mutation where the amino acid isoleucine becomes leucine, both of which are non-polar. However, figure 1b shows another mutation 1138D58 where large fragments of nucleotides are missing between positions 1138 G and 1158 C. This deletion is a frameshift mutation that shifts the grouping of the nucleotides, changing the code for the amino acids downstream from it. This could account for the inactivity of the tyrosinase due to incorrect amino acids being coded. Figure 1c has the 1467T68 mutation where an additional T nucleotide has been inserted into the TYR sequence. This addition also causes a frameshift mutation. Figure 1d shows the 127CT mutation where the change in nucleotides from C to T causes the codon normally for arginine to become a stop codon. This can affect TYR in that protein will be truncated. Figure 1e shows the 1150CG where C has changed to a G nucleotide. This missense mutation results in a change from histidine to asparagine. The histidine is basic and has a ring structure while asparagine is an amide. This drastic change will likely affect the overall structure of the tyrosinase protein. Finally, Figure 1f has 823GT where the expected G is instead a T due to a missense mutation. This causes the amino acid alanine to become a nucleophilic serine.
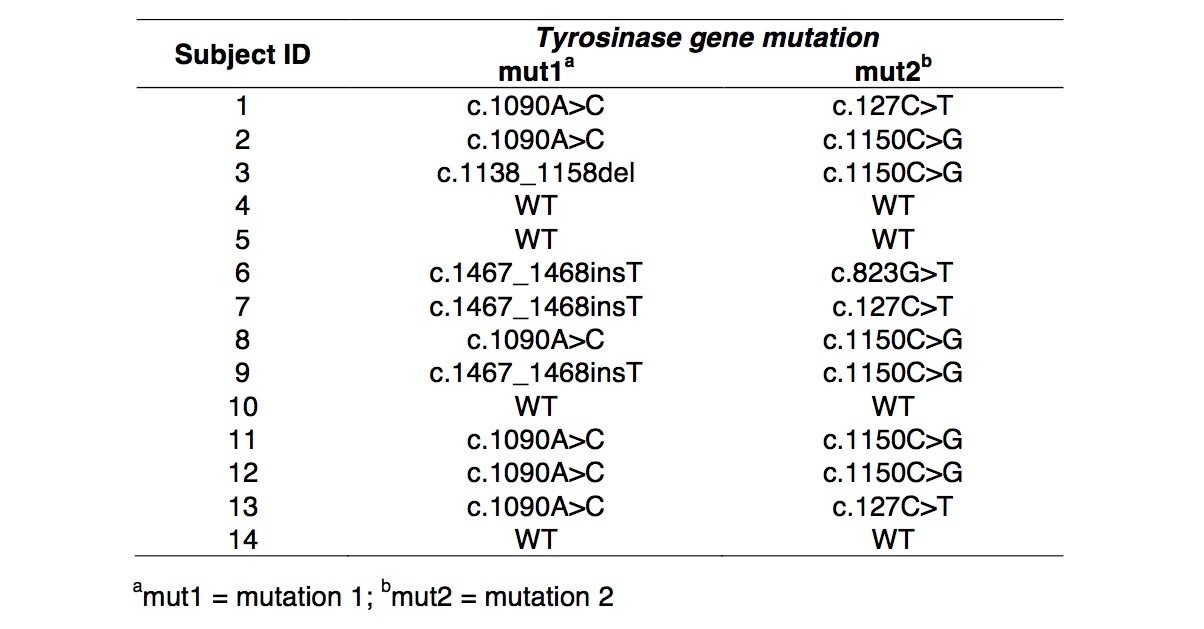 Table 3. (Click to Enlarge) TYR allele mutations.
Table 3. (Click to Enlarge) TYR allele mutations.
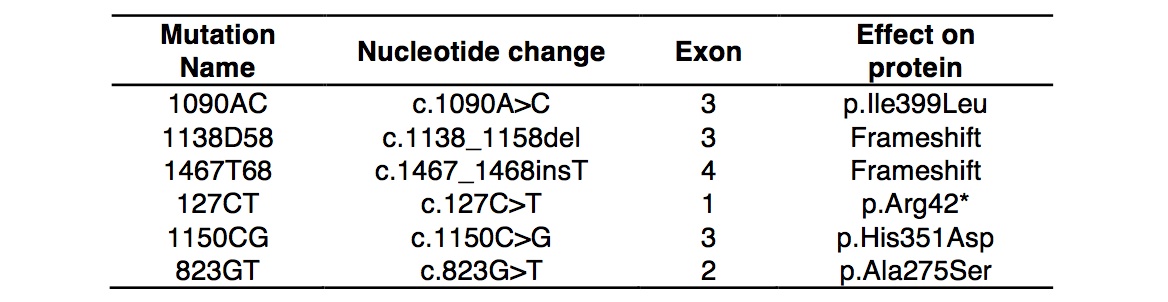 Table 4. (Click to Enlarge) Mutations in TYR gene and their effects on protein expression.
Table 4. (Click to Enlarge) Mutations in TYR gene and their effects on protein expression.
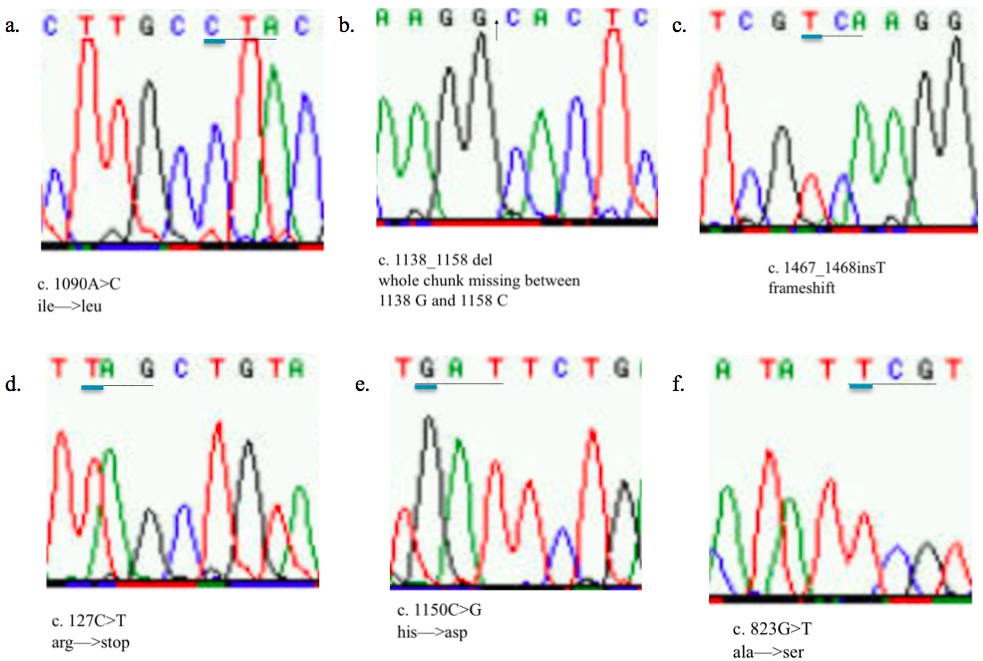 Figure 1. (Click to Enlarge) Direct sequence sections of the 6 mutations found in the tyrosinase gene.
Figure 1. (Click to Enlarge) Direct sequence sections of the 6 mutations found in the tyrosinase gene.
We examined the frequency of these 6 mutations from 10 OCA1 phenotypic Wookiees with our control group of 10 wild-type Wookiees and found that there were no mutations in the 20 alleles surveyed. Therefore, it is possible that the six novel mutations are pathologic and not polymorphic for OCA1. We also found that the missense mutations in TYR gene were the most common from our 10 TYR mutant OCA1 phenotype Wookiees, which is similar in humans [8].
In humans, the structure of tyrosinase contains 529 amino acids, with a signal peptide at the amino end and a transmembrane region at the carboxyl end [5]. The catalytic site of the enzyme contains two copper atoms ligated to six histidine residues. It also has glycosylation sites of asparagine residues at 86, 111, 161, 230, 384, and 499 [5]. The Wookiee TYR 1150CG mutation is located at the 384-residue site, which directly modifies one of these histidine amino acids and may have caused the inactivation of the mutated enzyme. The change in the sites of the asparagine residues would also have been affected by the Wookiee 1138D58 mutation since the nucleotides coding for the area were gone. In addition, the 1467T68 mutation is where there is a T nucleotide insertion changes all the amino acids downstream, thereby possibility affecting the site at 499. The 127CT mutation forms a stop codon relatively early, resulting in an incomplete tyrosinase protein that is unable to function normally. The 1090AC and 823GT mutations result in the change of isoleucine to leucine, and alanine to serene respectively. It is suspected that the change in amino acids caused an inactivation of the enzyme due to an alteration in the tertiary structure at the enzyme’s catalytic sites. This may also be a factor for the 1150CG, 1138D58, and 1467T68 mutations. Further studies will be required to verify why these six mutations cause a lack of tyrosinase activity.
Genotype-Phenotype Comparison
In humans, existing evidence from literature suggests that the genotype-phenotype correlation is strongest in individuals with two null TYR alleles [6]. However, attempts at genotype-phenotype correlations for OCA mutations have also proven difficult and have largely been unsuccessful because same TYR mutations are reported in patients with varying clinical presentations [6]. To date, more than 300 TYR variants have been reported to cause albinism in humans [6]. Furthermore, individuals who are heterozygous for the TYR mutant allele may demonstrate partial albinism wherein melanin is not completely absent and the affected individual is able to still produce some pigmentation. Conversely, our data suggest that partial albinism is potentially not present in Wookiees. As illustrated in Table 3, albinism in Wookiees is demonstrated when both TYR alleles are mutated. We did not observe any cases wherein an albino Wookiee was only heterozygous for the mutant TYR gene.
Previous studies have shown that OCA1 occurs in humans when the TYR activity is disrupted. Results of this study illustrate that this is also true for Wookiees (Table 3). Ten out of 14 albino Wookiee subjects have mutations on both alleles of their TYR genes. Interestingly, our data (Table 3) indicated that 4 of our OCA Wookiees with wild type TYR sequences also demonstrated albinism. Initial screening based on their phenotypic characteristics had suggested that these
albino Wookiees lacked tyrosinase activity, despite the lack of mutations in the TYR alleles. This finding is surprising and suggests that there may be other factors inhibiting the melanin production in Wookiees, by directly inhibiting tyrosinase activity, and that TYR is not the only gene that determines albinism in Wookiees. In humans, various studies have revealed that the mutation of several other genes gives rise to different types of albinism. It is possible that various melanogenic pathways may be present in Wookiees that have yet to be discovered, and that different modes of albinism may also exist.
There was a difference in the visual acuity in OCA Wookiees with the wild type TYR gene and the OCA Wookiees with the mutated TYR genes. Wookiees who expressed OCA1 but do not have a mutation in their TYR genes have a mean visual acuity of 0.75, whereas Wookiees with mutated TYR genes and OCA have a mean visual acuity of 0.27. This difference is statistically significant, having p
It is also interesting to note that the 4 albino Wookiees who had wild type TYR all came from the region of Korrokrrayyo, the tallest and most massive mountainous region in Kashyyyk (Table 5). It is impossible to conclude whether the region actually plays a role in the manifestation of albinism in Wookiees because of our limited knowledge regarding the environment and due to the small sample sized used in this study. However, it is important to acknowledge that environmental factors may certainly be critical in defining phenotypes, especially during early development. More studies would need to be conducted. Genes are not a steadfast blueprint for heredity. Specific environmental factors, which have yet to be determined, may be present in Korrokrrayyo that are affecting and altering gene expression in Wookiees.
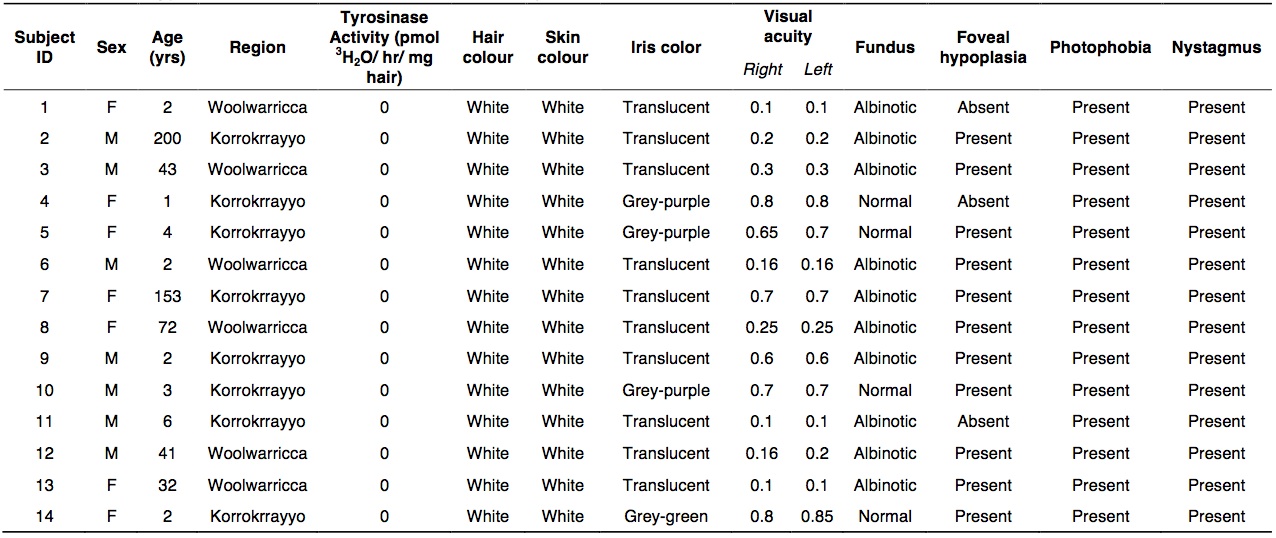 Table 5. (Click to Enlarge) Phenotypic characteristics of Wookies OCA1 subjects.
Table 5. (Click to Enlarge) Phenotypic characteristics of Wookies OCA1 subjects.
Acknowledgements
The authors thank the participants and their families for their participation. We also thank Mike Liverspokie for his technical assistance and Dr. Kankertop for critique of the manuscript. This work was supported in part by the Galactic Empire Research Foundation under Grant 226/2012-GERA
Literature Cited
1.National Aeronautics and Space Administration. Humanoid Life Form Discovered. Sci Int. 2006; 3: 23-30.
2. Hansen K, Damour H. Social Relationships: A Wookiee Perspective. J Psychol. 2010; 23: 887-911.
3. Holmes M, Rosenberg G, Ray Q. Using comparative genomics to evaluate evolutionary linkage between Wookiee Wookiees and Homo sapiens. Hum Genet. 2008; 2: 24-32.
4. Oetting W, King R. Molecular basis of albinism: mutations and polymorphisms of pigmentation genes associated with albinism. Hum Mutat. 1999; 13: 99-115
5. Nakamura E, Miyamura Y, Matsunaga J, Kano Y, Dakeishi-Hara M, Tanita M, Kono M, Tomita Y. A novel mutation of the tyrosinase causing oculocutaneous albinism type I (OCAI). J Dermatol Sci. 2002, 28: 102-5
6. Simeonov D, Wang X, Wang C, Sergeev Y, Dolinska M, Bower M, Fischer R, Winer D, Dubrovsky G, Balog J, Huizing M, Hart R, Zein W, Gahl W, Brooks B, Adams D. DNA Variations in oculocutaneous albinism: an updated mutation list and current outstanding issues in molecular diagnostics. Hum Mutat. 2013; 34: 827-35.
7. King R, Pietsch J, Fryer J, Savage S, Brott M, Russell-Eggitt I, Summers C, Oetting W. Tyrosinase gene mutations in oculocutaneous albinism 1 (OCA1): definition of the phenotype. Hum Genet. 2003; 113: 502-513.
8. Oetting W, Fryer J, Shriram S, King R. Oculocutaneous albinism type 1: the last 100 years. Pigment Cell Res. 2003; 16: 307-11.
9. Oetting W. The tyrosinase gene and oculocutaneous albinism type I (OCA1): a model for understanding the molecular biology of melanin formation. Pigment Cell Res. 2000; 13: 320-325.
10. Grønskov K, Ek J, Sand A, Scheller R, Bygum A, Brixen K, Brondum-Nielsen K, Rosenberg T. Birth prevalence and mutation spectrum in danish patients with autosomal recessive albinism. Invest Ophthalmol Vis Sci. 2009; 50: 1058-64.
11. Jaworek T, Kausar T, Bell S, Tariq N, Masgood M, Sohail A, Ali M, Igbal F, Rasool S, Shaikh R, Ahmed Z. Molecular genetic studies and delineation of the oculocutaneous albinism phenotype in the Pakistani population. Orphanet J Rare Dis. 2012; 7: 44
12. Tomita Y, Miyamura Y. Oculocutaneous albinism and analysis of tyrosinase gene in Japanese patients. Nagoya J Med Sci. 1998; 61: 97-102.
13. Shibahara S. Mutations of the tyrosinase gene in oculocutaneous albinism. Pigment Cell Res. 1992; 5: 279-83.